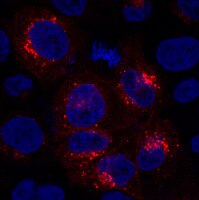
Induction and transport of Wnt 5a during macrophage-induced malignant invasion is mediated by two types of extracellular vesicles.
Menck, K; Klemm, F; Gross, JC; Pukrop, T; Wenzel, D; Binder, C
Oncotarget
4
2057-66
2013
Show Abstract
Recently, we have shown that macrophage (MΦ)-induced invasion of breast cancer cells requires upregulation of Wnt 5a in MΦ leading to activation of β-Catenin-independent Wnt signaling in the tumor cells. However, it remained unclear, how malignant cells induce Wnt 5a in MΦ and how it is transferred back to the cancer cells. Here we identify two types of extracellular particles as essential for this intercellular interaction in both directions. Plasma membrane-derived microvesicles (MV) as well as exosomes from breast cancer cells, although biologically distinct populations, both induce Wnt 5a in MΦ. In contrast, the particle-free supernatant and vesicles from benign cells, such as platelets, have no such effect. Induction is antagonized by the Wnt inhibitor Dickkopf-1. Subsequently, Wnt 5a is shuttled via responding MΦ-MV and exosomes to the tumor cells enhancing their invasion. Wnt 5a export on both vesicle fractions depends at least partially on the cargo protein Evenness interrupted (Evi). Its knockdown leads to Wnt 5a depletion of both particle populations and reduced vesicle-mediated invasion. In conclusion, MV and exosomes are critical for MΦ-induced invasion of cancer cells since they are responsible for upregulation of MΦ-Wnt 5a as well as for its delivery to the recipient cells via a reciprocal loop. Although of different biogenesis, both populations share common features regarding function and Evi-dependent secretion of non-canonical Wnts. | 24185202
 |
A uniform human Wnt expression library reveals a shared secretory pathway and unique signaling activities.
Najdi, R; Proffitt, K; Sprowl, S; Kaur, S; Yu, J; Covey, TM; Virshup, DM; Waterman, ML
Differentiation; research in biological diversity
84
203-13
2011
Show Abstract
Wnt ligands are secreted morphogens that control multiple developmental processes during embryogenesis and adult homeostasis. A diverse set of receptors and signals have been linked to individual Wnts, but the lack of tools for comparative analysis has limited the ability to determine which of these signals are general for the entire Wnt family, and which define subsets of differently acting ligands. We have created a versatile Gateway library of clones for all 19 human Wnts. An analysis comparing epitope-tagged and untagged versions of each ligand shows that despite their similar expression at the mRNA level, Wnts exhibit considerable variation in stability, processing and secretion. At least 14 out of the 19 Wnts activate β-catenin-dependent signaling, an activity that is cell type-dependent and tracks with the stabilization of β-catenin and LRP6 phosphorylation. We find that the core Wnt modification and secretion proteins Porcupine (PORCN) and Wntless (WLS) are essential for all Wnts to signal through β-catenin-dependent and independent pathways. This comprehensive toolkit provides critical tools and new insights into human Wnt gene expression and function. | 22784633
|
WLS-dependent secretion of WNT3A requires Ser209 acylation and vacuolar acidification.
Coombs, Gary S, et al.
J. Cell. Sci., 123: 3357-67 (2010)
2009
Show Abstract
Wnt proteins are secreted post-translationally modified proteins that signal locally to regulate development and proliferation. The production of bioactive Wnts requires a number of dedicated factors in the secreting cell whose coordinated functions are not fully understood. A screen for small molecules identified inhibitors of vacuolar acidification as potent inhibitors of Wnt secretion. Inhibition of the V-ATPase or disruption of vacuolar pH gradients by diverse drugs potently inhibited Wnt/β-catenin signaling both in cultured human cells and in vivo, and impaired Wnt-regulated convergent extension movements in Xenopus embryos. WNT secretion requires its binding to the carrier protein wntless (WLS); we find that WLS is ER-resident in human cells and WNT3A binding to WLS requires PORCN-dependent lipid modification of WNT3A at serine 209. Inhibition of vacuolar acidification results in accumulation of the WNT3A-WLS complex both in cells and at the plasma membrane. Modeling predictions suggest that WLS has a lipid-binding β-barrel that is similar to the lipocalin-family fold. We propose that WLS binds Wnts in part through a lipid-binding domain, and that vacuolar acidification is required to release palmitoylated WNT3A from WLS in secretory vesicles, possibly to facilitate transfer of WNT3A to a soluble carrier protein. | 20826466
|
Reciprocal regulation of Wnt and Gpr177/mouse Wntless is required for embryonic axis formation.
Fu, Jiang, et al.
Proc. Natl. Acad. Sci. U.S.A., 106: 18598-603 (2009)
2009
Show Abstract
Members of the Wnt family are secreted glycoproteins that trigger cellular signals essential for proper development of organisms. Cellular signaling induced by Wnt proteins is involved in diverse developmental processes and human diseases. Previous studies have generated an enormous wealth of knowledge on the events in signal-receiving cells. However, relatively little is known about the making of Wnt in signal-producing cells. Here, we describe that Gpr177, the mouse orthologue of Drosophila Wls, is expressed during formation of embryonic axes. Embryos with deficient Gpr177 exhibit defects in establishment of the body axis, a phenotype highly reminiscent to the loss of Wnt3. Although many different mammalian Wnt proteins are required for a wide range of developmental processes, the Wnt3 ablation exhibits the earliest developmental abnormality. This suggests that the Gpr177-mediated Wnt production cannot be substituted. As a direct target of Wnt, Gpr177 is activated by beta-catenin and LEF/TCF-dependent transcription. This activation alters the cellular distributions of Gpr177 which binds to Wnt proteins and assists their sorting and secretion in a feedback regulatory mechanism. Our findings demonstrate that the loss of Gpr177 affects Wnt production in the signal-producing cells, leading to alterations of Wnt signaling in the signal-receiving cells. A reciprocal regulation of Wnt and Gpr177 is essential for the patterning of the anterior-posterior axis during mammalian development. | 19841259
|
Wntless, a conserved membrane protein dedicated to the secretion of Wnt proteins from signaling cells.
Bänziger, Carla, et al.
Cell, 125: 509-22 (2006)
2005
Show Abstract
Cell-cell communication via Wnt signals represents a fundamental means by which animal development and homeostasis are controlled. The identification of components of the Wnt pathway is reaching saturation for the transduction process in receiving cells but is incomplete concerning the events occurring in Wnt-secreting cells. Here, we describe the discovery of a novel Wnt pathway component, Wntless (Wls/Evi), and show that it is required for Wingless-dependent patterning processes in Drosophila, for MOM-2-governed polarization of blastomeres in C. elegans, and for Wnt3a-mediated communication between cultured human cells. In each of these cases, Wls is acting in the Wnt-sending cells to promote the secretion of Wnt proteins. Since loss of Wls function has no effect on other signaling pathways yet appears to impede all the Wnt signals we analyzed, we propose that Wls represents an ancient partner for Wnts dedicated to promoting their secretion into the extracellular milieu. | 16678095
|