Wenn Sie das Fenster schließen, wird Ihre Konfiguration nicht gespeichert, es sei denn, Sie haben Ihren Artikel in die Bestellung aufgenommen oder zu Ihren Favoriten hinzugefügt.
Klicken Sie auf OK, um das MILLIPLEX® MAP-Tool zu schließen oder auf Abbrechen, um zu Ihrer Auswahl zurückzukehren.
Wählen Sie konfigurierbare Panels & Premixed-Kits - ODER - Kits für die zelluläre Signaltransduktion & MAPmates™
Konfigurieren Sie Ihre MILLIPLEX® MAP-Kits und lassen sich den Preis anzeigen.
Konfigurierbare Panels & Premixed-Kits
Unser breites Angebot enthält Multiplex-Panels, für die Sie die Analyten auswählen können, die am besten für Ihre Anwendung geeignet sind. Unter einem separaten Register können Sie das Premixed-Cytokin-Format oder ein Singleplex-Kit wählen.
Kits für die zelluläre Signaltransduktion & MAPmates™
Wählen Sie gebrauchsfertige Kits zur Erforschung gesamter Signalwege oder Prozesse. Oder konfigurieren Sie Ihre eigenen Kits mit Singleplex MAPmates™.
Die folgenden MAPmates™ sollten nicht zusammen analysiert werden: -MAPmates™, die einen unterschiedlichen Assaypuffer erfordern. -Phosphospezifische und MAPmate™ Gesamtkombinationen wie Gesamt-GSK3β und Gesamt-GSK3β (Ser 9). -PanTyr und locusspezifische MAPmates™, z.B. Phospho-EGF-Rezeptor und Phospho-STAT1 (Tyr701). -Mehr als 1 Phospho-MAPmate™ für ein einziges Target (Akt, STAT3). -GAPDH und β-Tubulin können nicht mit Kits oder MAPmates™, die panTyr enthalten, analysiert werden.
.
Bestellnummer
Bestellinformationen
St./Pkg.
Liste
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Wählen Sie bitte Spezies, Panelart, Kit oder Probenart
Um Ihr MILLIPLEX® MAP-Kit zu konfigurieren, wählen Sie zunächst eine Spezies, eine Panelart und/oder ein Kit.
Custom Premix Selecting "Custom Premix" option means that all of the beads you have chosen will be premixed in manufacturing before the kit is sent to you.
Catalogue Number
Ordering Description
Qty/Pack
List
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Spezies
Panelart
Gewähltes Kit
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
96-Well Plate
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
Weitere Reagenzien hinzufügen (MAPmates erfordern die Verwendung eines Puffer- und Detektionskits)
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
48-602MAG
Buffer Detection Kit for Magnetic Beads
1 Kit
Platzsparende Option Kunden, die mehrere Kits kaufen, können ihre Multiplex-Assaykomponenten in Kunststoffbeuteln anstelle von Packungen erhalten, um eine kompaktere Lagerung zu ermöglichen.
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Das Produkt wurde in Ihre Bestellung aufgenommen
Sie können nun ein weiteres Kit konfigurieren, ein Premixed-Kit wählen, zur Kasse gehen oder das Bestell-Tool schließen.
06-270
Sigma-AldrichAnti-Giα3 Antibody
Detect Giα3 using this Anti-Giα3 Antibody validated for use in IP & WB.
More>>Detect Giα3 using this Anti-Giα3 Antibody validated for use in IP & WB. Less<<
Anti-Giα3 Antibody: SDB (Sicherheitsdatenblätter), Analysenzertifikate und Qualitätszertifikate, Dossiers, Broschüren und andere verfügbare Dokumente.
FUNCTION: SwissProt: P08754 # Guanine nucleotide-binding proteins (G proteins) are involved as modulators or transducers in various transmembrane signaling systems. G(k) is the stimulatory G protein of receptor- regulated K(+) channels. SIZE: 354 amino acids; 40532 Da SUBUNIT: G proteins are composed of 3 units; alpha, beta and gamma. The alpha chain contains the guanine nucleotide binding site. Interacts with GPSM1 (By similarity). SIMILARITY: SwissProt: P08754 ## Belongs to the G-alpha family. G(i/o/t/z) subfamily.
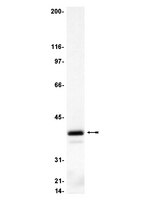
Molecular Weight
40 kDa
Physicochemical Information
Dimensions
Materials Information
Toxicological Information
Safety Information according to GHS
Safety Information
Product Usage Statements
Quality Assurance
routinely evaluated by immunoblot on rat hippocampus membrane preparation, solubilized rat and mouse brain membranes or mouse 3T3/A31 cell lysates
Usage Statement
Unless otherwise stated in our catalog or other company documentation accompanying the product(s), our products are intended for research use only and are not to be used for any other purpose, which includes but is not limited to, unauthorized commercial uses, in vitro diagnostic uses, ex vivo or in vivo therapeutic uses or any type of consumption or application to humans or animals.
Storage and Shipping Information
Storage Conditions
Maintain for 2 years at -20°C from date of shipment. Aliquot to avoid repeated freezing and thawing. For maximum recovery of product, centrifuge the original vial after thawing and prior to removing the cap.
Subcellular localization of LGN during mitosis: evidence for its cortical localization in mitotic cell culture systems and its requirement for normal cell cycle progression. Kaushik, R; Yu, F; Chia, W; Yang, X; Bahri, S Molecular biology of the cell
14
3144-55
2003
Mammalian LGN/AGS3 proteins and their Drosophila Pins orthologue are cytoplasmic regulators of G-protein signaling. In Drosophila, Pins localizes to the lateral cortex of polarized epithelial cells and to the apical cortex of neuroblasts where it plays important roles in their asymmetric division. Using overexpression studies in different cell line systems, we demonstrate here that, like Drosophila Pins, LGN can exhibit enriched localization at the cell cortex, depending on the cell cycle and the culture system used. We find that in WISH, PC12, and NRK but not COS cells, LGN is largely directed to the cell cortex during mitosis. Overexpression of truncated protein domains further identified the Galpha-binding C-terminal portion of LGN as a sufficient domain for cortical localization in cell culture. In mitotic COS cells that normally do not exhibit cortical LGN localization, LGN is redirected to the cell cortex upon overexpression of Galpha subunits of heterotrimeric G-proteins. The results also show that the cortical localization of LGN is dependent on microfilaments and that interfering with LGN function in cultured cell lines causes early disruption to cell cycle progression.
A role for tyrosine phosphorylation in the regulation and sensitization of adenylate cyclase by melatonin Barrett, P., et al Faseb J, 14:1619-28 (2000)
1999
Disulfide bonds in the extracellular calcium-polyvalent cation-sensing receptor correlate with dimer formation and its response to divalent cations in vitro Ward, D. T., et al J Biol Chem, 273:14476-83 (1998)
1998
Apical extracellular calcium/polyvalent cation-sensing receptor regulates vasopressin-elicited water permeability in rat kidney inner medullary collecting duct Sands, J. M., et al J Clin Invest, 99:1399-405 (1997)
1997
G(o)-protein alpha-subunits activate mitogen-activated protein kinase via a novel protein kinase C-dependent mechanism. van Biesen, T, et al. J. Biol. Chem., 271: 1266-9 (1996)
1996
Mitogen-activated protein kinase (MAPK) is activated in response to both receptor tyrosine kinases and G-protein-coupled receptors. Recently, Gi-coupled receptors, such as the alpha 2A adrenergic receptor, were shown to mediate Ras-dependent MAPK activation via a pathway requiring G-protein beta gamma subunits (G beta gamma) and many of the same intermediates involved in receptor tyrosine kinase signaling. In contrast, Gq-coupled receptors, such as the M1 muscarinic acetylcholine receptor (M1AChR), activate MAPK via a pathway that is Ras-independent but requires the activity of protein kinase C (PKC). Here we show that, in Chinese hamster ovary cells, the M1AChR and platelet-activating factor receptor (PAFR) mediate MAPK activation via the alpha-subunit of the G(o) protein. G(o)-mediated MAPK activation was sensitive to treatment with pertussis toxin but insensitive to inhibition by a G beta gamma-sequestering peptide (beta ARK1ct). M1AChR and PAFR catalyzed G(o) alpha-subunit GTP exchange, and MAPK activation could be partially rescued by a pertussis toxin-insensitive mutant of G(o) alpha but not by similar mutants of Gi. G(o)-mediated MAPK activation was insensitive to inhibition by a dominant negative mutant of Ras (N17Ras) but was completely blocked by cellular depletion of PKC. Thus, M1AChR and PAFR, which have previously been shown to couple to Gq, are also coupled to G(o) to activate a novel PKC-dependent mitogenic signaling pathway.
Guanine nucleotide binding regulatory proteins and adenylate cyclase in livers of streptozotocin- and BB/Wor-diabetic rats. Immunodetection of Gs and Gi with antisera prepared against synthetic peptides. Lynch, C J, et al. J. Clin. Invest., 83: 2050-62 (1989)
1988
Adenylate cyclase in liver plasma membranes from streptozotocin-diabetic (STZ) or BB/Wor spontaneously diabetic rats showed increased responsiveness to GTP, glucagon, fluoroaluminate, and cholera toxin. Basal or forskolin-stimulated activity was unchanged in STZ rats, but increased in BB/Wor rats. No change in the alpha-subunit of Gi (alpha i) was observed in STZ or BB/Wor rats using pertussis toxin-stimulated [32P]ADP-ribosylation. Immunodetection using antibodies against the COOH-terminal decapeptides of alpha T and alpha i-3 showed no change in alpha i in STZ rats and a slight decrease in BB/Wor rats. Angiotensin II inhibition of hepatic adenylate cyclase was not altered in either diabetic rat. In both models of diabetes, Gs alpha-subunits were increased as measured by cholera toxin-stimulated [32P]-ADP-ribosylation of 43-47.5-kD peptides, reconstitution with membranes from S49 cyc- cells or immunoreactivity using antibodies against the COOH-terminal decapeptide of alpha s. These data indicate that STZ-diabetes increases hepatic Gs but does not change Gi or adenylate cyclase catalytic activity. In contrast, BB/Wor rats show increased hepatic Gs and adenylate cyclase. These changes could explain the increase in hepatic cAMP and related dysfunctions observed in diabetes.