Wenn Sie das Fenster schließen, wird Ihre Konfiguration nicht gespeichert, es sei denn, Sie haben Ihren Artikel in die Bestellung aufgenommen oder zu Ihren Favoriten hinzugefügt.
Klicken Sie auf OK, um das MILLIPLEX® MAP-Tool zu schließen oder auf Abbrechen, um zu Ihrer Auswahl zurückzukehren.
Wählen Sie konfigurierbare Panels & Premixed-Kits - ODER - Kits für die zelluläre Signaltransduktion & MAPmates™
Konfigurieren Sie Ihre MILLIPLEX® MAP-Kits und lassen sich den Preis anzeigen.
Konfigurierbare Panels & Premixed-Kits
Unser breites Angebot enthält Multiplex-Panels, für die Sie die Analyten auswählen können, die am besten für Ihre Anwendung geeignet sind. Unter einem separaten Register können Sie das Premixed-Cytokin-Format oder ein Singleplex-Kit wählen.
Kits für die zelluläre Signaltransduktion & MAPmates™
Wählen Sie gebrauchsfertige Kits zur Erforschung gesamter Signalwege oder Prozesse. Oder konfigurieren Sie Ihre eigenen Kits mit Singleplex MAPmates™.
Die folgenden MAPmates™ sollten nicht zusammen analysiert werden: -MAPmates™, die einen unterschiedlichen Assaypuffer erfordern. -Phosphospezifische und MAPmate™ Gesamtkombinationen wie Gesamt-GSK3β und Gesamt-GSK3β (Ser 9). -PanTyr und locusspezifische MAPmates™, z.B. Phospho-EGF-Rezeptor und Phospho-STAT1 (Tyr701). -Mehr als 1 Phospho-MAPmate™ für ein einziges Target (Akt, STAT3). -GAPDH und β-Tubulin können nicht mit Kits oder MAPmates™, die panTyr enthalten, analysiert werden.
.
Bestellnummer
Bestellinformationen
St./Pkg.
Liste
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Wählen Sie bitte Spezies, Panelart, Kit oder Probenart
Um Ihr MILLIPLEX® MAP-Kit zu konfigurieren, wählen Sie zunächst eine Spezies, eine Panelart und/oder ein Kit.
Custom Premix Selecting "Custom Premix" option means that all of the beads you have chosen will be premixed in manufacturing before the kit is sent to you.
Catalogue Number
Ordering Description
Qty/Pack
List
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Spezies
Panelart
Gewähltes Kit
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
96-Well Plate
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
Weitere Reagenzien hinzufügen (MAPmates erfordern die Verwendung eines Puffer- und Detektionskits)
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
48-602MAG
Buffer Detection Kit for Magnetic Beads
1 Kit
Platzsparende Option Kunden, die mehrere Kits kaufen, können ihre Multiplex-Assaykomponenten in Kunststoffbeuteln anstelle von Packungen erhalten, um eine kompaktere Lagerung zu ermöglichen.
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Das Produkt wurde in Ihre Bestellung aufgenommen
Sie können nun ein weiteres Kit konfigurieren, ein Premixed-Kit wählen, zur Kasse gehen oder das Bestell-Tool schließen.
Anti-Cytokeratin 8/18, clone L2A1, Cat. No. MABT844, is a mouse monoclonal antibody that detects cytokeratin 8/18 and has been tested for use in Immunocytochemistry and Immunoprecipitation.
More>>Anti-Cytokeratin 8/18, clone L2A1, Cat. No. MABT844, is a mouse monoclonal antibody that detects cytokeratin 8/18 and has been tested for use in Immunocytochemistry and Immunoprecipitation. Less<<
Keratin type II cytoskeletal 8/Keratin type I cytoskeletal 18
Keratin-8/Keratin-18
K8/K18)
Background Information
Cytokeratin-8/18 (UniProt: P05787/P05783; also known as Keratin, type II cytoskeletal 8/ Keratin, type I cytoskeletal 18, Keratin-8/Keratin-18, K8/K18) is encoded by the KRT8/KRT18 (also known as CYK8/CYK18) gene (Gene ID: 3856/3875) in human. Cytokeratins 8 and 18 (K8/18) are simple epithelial cell-specific intermediate filament proteins. Cytokeratin-8 is a type II, neutral to basic, protein of the intermediate filament family that together with Cytokeratin-19 (KRT19) helps to link the contractile apparatus to dystrophin at the costameres of striated muscle. Cytokeratin-8 can undergo phosphorylation on three major serine residues: Serine 23, 431, and 73. Serine 23 is shown to be highly conserved in all type II keratins. Phosphorylation at Serine 73 is reported to increase during cellular stress, including hear and drug exposure. However, under normal conditions serine 73 remains largely dephosphorylated. Cytokeratin-8 can also undergo O-glycosylation in a cell cycle-dependent manner and glycosylation increases its solubility and reduces stability by inducing proteasomal degradation. Cytokeratin-18 is a type I, acidic, protein that is expressed in colon, placenta, and liver. Higher expression levels have been reported in lymph nodes of breast carcinoma. Cytokeratin-18 is involved in the uptake of thrombin-antithrombin complexes by hepatic cells. Upon phosphorylation, it plays a role in filament reorganization. It is proteolytically cleaved by caspases during epithelial cell apoptosis and this cleavage is shown to occur at Asp-238. Phosphorylation of cytokeratin-18 at serine 33 is shown to increase during mitosis in cultured cells and in regenerating liver. Serine 33 phosphorylation is considered to be essential for its association with 14-3-3 proteins and has a role in keratin organization and distribution. Cytokeratin-18 can undergo O-glycosylation, which results in its increased solubility and reduction in its stability. Mutations in KRT8 and KRT18 genes have been been linked to liver cirrhosis that is characterized by severe panlobular liver-cell swelling with Mallory body formation, prominent pericellular fibrosis, and marked deposits of copper.
References
Product Information
Format
Purified
Presentation
Purified mouse monoclonal antibody IgG in buffer containing 0.1 M Tris-Glycine (pH 7.4), 150 mM NaCl with 0.05% sodium azide.
Applications
Application
Anti-Cytokeratin 8/18, clone L2A1, Cat. No. MABT844, is a mouse monoclonal antibody that detects cytokeratin 8/18 and has been tested for use in Immunocytochemistry and Immunoprecipitation.
Key Applications
Immunocytochemistry
Immunoprecipitation
Application Notes
Immunoprecipitation Analysis: A representative lot detected Cytokeratin 8/18 in Immunoprecipitation applications (Haltiwanger, R.S., et. al. (1997). J Biol Chem. 272(13):8752-8; Chou, C.F., et. al. (1994). Biochem J. 298 ( Pt 2):457-63; Chou, C.F., et. al. (1993). Biochem J. 268(6):4465-72; Ku, N.O., et. al. (2000). J Cell Biol. 149(3):547-52; Lahdeniemi, I.A.K., et. al. (2017). Cell Death Differ. 24(6):984-996).
Immunocytochemistry Analysis: A representative lot detected Cytokeratin 8/18 in Immunocytochemistry applications (Chou, C.F., et. al. (1993). Biochem J. 268(6):4465-72).
Biological Information
Immunogen
KLH-conjugated Linear Peptide corresponding to a sequence from Human Cytokeratin 8/18.
Clone
L2A1
Concentration
Please refer to lot specific datasheet.
Host
Mouse
Specificity
Clone L2A1 is a mouse monoclonal antibody that detects human cytokeratin 8/18.
~53.70 kDa and 48.06 kDa, respectively for cytokeratin 8 and 18 calculated. Uncharacterized bands may be observed in some lysate(s).
Physicochemical Information
Dimensions
Materials Information
Toxicological Information
Safety Information according to GHS
Safety Information
Product Usage Statements
Quality Assurance
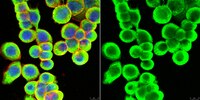
Evaluated by Immunocytochemistry in HT-29 cells.
Immunocytochemistry Analysis: A 1:500 dilution of this antibody detected Cytokeratin 8/18 in HT-29 cells.
Usage Statement
Unless otherwise stated in our catalog or other company documentation accompanying the product(s), our products are intended for research use only and are not to be used for any other purpose, which includes but is not limited to, unauthorized commercial uses, in vitro diagnostic uses, ex vivo or in vivo therapeutic uses or any type of consumption or application to humans or animals.
Regnase-1 Maintains Iron Homeostasis via the Degradation of Transferrin Receptor 1 and Prolyl-Hydroxylase-Domain-Containing Protein 3 mRNAs. Yoshinaga, M; Nakatsuka, Y; Vandenbon, A; Ori, D; Uehata, T; Tsujimura, T; Suzuki, Y; Mino, T; Takeuchi, O Cell Rep
19
1614-1630
2016
Iron metabolism is regulated by transcriptional and post-transcriptional mechanisms. The mRNA of the iron-controlling gene, transferrin receptor 1 (TfR1), has long been believed to be negatively regulated by a yet-unidentified endonuclease. Here, we show that the endonuclease Regnase-1 is critical for the degradation of mRNAs involved in iron metabolism in vivo. First, we demonstrate that Regnase-1 promotes TfR1 mRNA decay. Next, we show that Regnase-1-/- mice suffer from severe iron deficiency anemia, although hepcidin expression is downregulated. The iron deficiency anemia is induced by a defect in duodenal iron uptake. We reveal that duodenal Regnase-1 controls the expression of PHD3, which impairs duodenal iron uptake via HIF2α suppression. Finally, we show that Regnase-1 is a HIF2α-inducible gene and thus provides a positive feedback loop for HIF2α activation via PHD3. Collectively, these results demonstrate that Regnase-1-mediated regulation of iron-related transcripts is essential for the maintenance of iron homeostasis.
Keratin polypeptides 8 and 18 (K8/18) are intermediate filament (IF) proteins that are expressed in glandular epithelia. Although the mechanism of keratin turnover is poorly understood, caspase-mediated degradation of type I keratins occurs during apoptosis and the proteasome pathway has been indirectly implicated in keratin turnover based on colocalization of keratin-ubiquitin antibody staining. Here we show that K8 and K18 are ubiquitinated based on cotransfection of His-tagged ubiquitin and human K8 and/or K18 cDNAs, followed by purification of ubiquitinated proteins and immunoblotting with keratin antibodies. Transfection of K8 or K18 alone yields higher levels of keratin ubiquitination as compared with cotransfection of K8/18, likely due to stabilization of the keratin heteropolymer. Most of the ubiquitinated species partition with the noncytosolic keratin fraction. Proteasome inhibition stabilizes K8 and K18 turnover, and is associated with accumulation of phosphorylated keratins, which indicates that although keratins are stable they still turnover. Analysis of K8 and K18 ubiquitination and degradation showed that K8 phosphorylation contributes to its stabilization. Our results provide direct evidence for K8 and K18 ubiquitination, in a phosphorylation modulated fashion, as a mechanism for regulating their turnover and suggest that other IF proteins could undergo similar regulation. These and other data offer a model that links keratin ubiquitination and hyperphosphorylation that, in turn, are associated with Mallory body deposits in a variety of liver diseases.
Mitotic arrest with nocodazole induces selective changes in the level of O-linked N-acetylglucosamine and accumulation of incompletely processed N-glycans on proteins from HT29 cells. Haltiwanger, RS; Philipsberg, GA J Biol Chem
272
8752-8
1997
O-Linked N-acetylglucosamine (O-GlcNAc) is a ubiquitous and abundant protein modification found on nuclear and cytoplasmic proteins. Several lines of evidence suggest that it is a highly dynamic modification and that the levels of this sugar on proteins may be regulated. Previous workers (Chou, C. F., and Omary, M. B. (1993) J. Biol. Chem. 268, 4465-4472) have shown that mitotic arrest with microtubule-destabilizing agents such as nocodazole causes an increase in the O-GlcNAc levels on keratins in the human colon cancer cell line HT29. We have sought to determine whether this increase in glycosylation is a general (i.e. occurring on many proteins) or a limited (i.e. occurring only on the keratins) process. A general increase would suggest that the microtubule-destabilizing agents were somehow affecting the enzymes responsible for addition and/or removal of O-GlcNAc. Our results suggest that the changes in O-GlcNAc induced by nocodazole are selective for the keratins. The levels of O-GlcNAc on other proteins, including the nuclear pore protein p62 and the transcription factor Sp1, are not significantly affected by this treatment. In agreement with these findings, nocodazole treatment caused no change in the activity of the enzymes responsible for addition or removal of O-GlcNAc as determined by direct in vitro assay. Interestingly, nocodazole treatment did cause a dramatic increase in modification of N-glycans with terminal GlcNAc residues on numerous proteins. Potential mechanisms for this and the change in glycosylation of the keratins are discussed.
Identification of a keratin-associated protein that localizes to a membrane compartment. Chou, CF; Riopel, CL; Omary, MB Biochem J
298 ( Pt 2)
457-63
1993
We describe the characterization of an acidic glycoprotein (molecular mass approximately 85 kDa) that associates with keratin intermediate filaments of 'simple'-type epithelia. Using a number of anti-keratin monoclonal antibodies, the 85 kDa glycoprotein was identified by co-immunoprecipitation with keratin polypeptides 8 and 18 (K8/18) from the human colonic epithelial cell line HT29 and several other epithelial cell lines. This Keratin-Associated Protein (termed KAP85) was readily detected after in vitro galactosylation of K8/18 immunoprecipitates obtained from mitosis-arrested cells. Its solubilization and detection were dependent on the detergent used, and it was barely detected after in vitro galactosylation of asynchronously growing G0/G1-phase cells. Its poor in vitro galactosylation in G0/G1-phase cells is likely a reflection of the lack of available terminal N-acetylglucosamine residues, since it can be labelled to a similar extent in G0/G1- and G2/M-phase cells using NaIO4/NaB3H4. Glycosidase digestion showed that KAP85 contains high mannose and complex oligosaccharides. Fractionation of total cellular K8/18 into soluble and cytoskeletal insoluble pools showed that KAP85 associates exclusively with the cytoskeletal K8/18 pool. Subcellular fractionation showed that KAP85 co-localizes with a plasma-membrane-enriched fraction that includes the transferrin receptor and KS-1 antigen. Our results demonstrate in vitro evidence of a membrane-associated glycoprotein (KAP85) which may serve as an attachment site for filamentous K8/18.
Mitotic arrest-associated enhancement of O-linked glycosylation and phosphorylation of human keratins 8 and 18. Chou, CF; Omary, MB J Biol Chem
268
4465-72
1992
Arrest of the human colonic cell line HT29 at the G2/M phase of the cell cycle resulted in changes in keratin assembly that were coupled with a significant increase in the O-linked glycosylation and serine phosphorylation of keratin polypeptides 8 and 18 (K8/18). With mitotic arrest, enhanced keratin phosphorylation occurred preferentially on K8, whereas K18 showed a higher glycosylation level than K8. Removal of the arresting agent allowed cells to proceed through the cell cycle with a concomitant decrease in K8/18 glycosylation. In contrast, keratins isolated from S phase-enriched cells, obtained after synchronization with aphidicolin, did not show enhanced glycosylation. Tryptic peptide analysis of keratins in G2/M-arrested cells showed changes in the glycopeptide pattern of K8 and in the phosphopeptide patterns of K8 and K18. Labeling of K8/18 immunoprecipitates, isolated from G2/M-arrested cells, with [3H]galactose followed by beta-elimination showed that K8/18 glycosylation consisted of single N-acetylglucosamine residues. Threonine was identified as the site of glycosylation after comparing acid hydrolysis products of beta-eliminated and non-beta-eliminated K8 and K18. Specific cleavage at tryptophan residues indicated that K18 glycosylation and phosphorylation were restricted to the head and proximal rod domains, whereas K8 did not show the same restriction. Our results show a unique association of the single O-linked N-acetylglucosamine type of modification of keratins with mitotic arrest in HT29 cells. There was no reciprocal relationship between K8/18 glycosylation and phosphorylation, and each keratin showed a preferential G2/M cell cycle-associated increase in either serine phosphorylation or threonine glycosylation.