Wenn Sie das Fenster schließen, wird Ihre Konfiguration nicht gespeichert, es sei denn, Sie haben Ihren Artikel in die Bestellung aufgenommen oder zu Ihren Favoriten hinzugefügt.
Klicken Sie auf OK, um das MILLIPLEX® MAP-Tool zu schließen oder auf Abbrechen, um zu Ihrer Auswahl zurückzukehren.
Wählen Sie konfigurierbare Panels & Premixed-Kits - ODER - Kits für die zelluläre Signaltransduktion & MAPmates™
Konfigurieren Sie Ihre MILLIPLEX® MAP-Kits und lassen sich den Preis anzeigen.
Konfigurierbare Panels & Premixed-Kits
Unser breites Angebot enthält Multiplex-Panels, für die Sie die Analyten auswählen können, die am besten für Ihre Anwendung geeignet sind. Unter einem separaten Register können Sie das Premixed-Cytokin-Format oder ein Singleplex-Kit wählen.
Kits für die zelluläre Signaltransduktion & MAPmates™
Wählen Sie gebrauchsfertige Kits zur Erforschung gesamter Signalwege oder Prozesse. Oder konfigurieren Sie Ihre eigenen Kits mit Singleplex MAPmates™.
Die folgenden MAPmates™ sollten nicht zusammen analysiert werden: -MAPmates™, die einen unterschiedlichen Assaypuffer erfordern. -Phosphospezifische und MAPmate™ Gesamtkombinationen wie Gesamt-GSK3β und Gesamt-GSK3β (Ser 9). -PanTyr und locusspezifische MAPmates™, z.B. Phospho-EGF-Rezeptor und Phospho-STAT1 (Tyr701). -Mehr als 1 Phospho-MAPmate™ für ein einziges Target (Akt, STAT3). -GAPDH und β-Tubulin können nicht mit Kits oder MAPmates™, die panTyr enthalten, analysiert werden.
.
Bestellnummer
Bestellinformationen
St./Pkg.
Liste
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Wählen Sie bitte Spezies, Panelart, Kit oder Probenart
Um Ihr MILLIPLEX® MAP-Kit zu konfigurieren, wählen Sie zunächst eine Spezies, eine Panelart und/oder ein Kit.
Custom Premix Selecting "Custom Premix" option means that all of the beads you have chosen will be premixed in manufacturing before the kit is sent to you.
Catalogue Number
Ordering Description
Qty/Pack
List
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Spezies
Panelart
Gewähltes Kit
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
96-Well Plate
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
Weitere Reagenzien hinzufügen (MAPmates erfordern die Verwendung eines Puffer- und Detektionskits)
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
48-602MAG
Buffer Detection Kit for Magnetic Beads
1 Kit
Platzsparende Option Kunden, die mehrere Kits kaufen, können ihre Multiplex-Assaykomponenten in Kunststoffbeuteln anstelle von Packungen erhalten, um eine kompaktere Lagerung zu ermöglichen.
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Das Produkt wurde in Ihre Bestellung aufgenommen
Sie können nun ein weiteres Kit konfigurieren, ein Premixed-Kit wählen, zur Kasse gehen oder das Bestell-Tool schließen.
Anti-Thrombomodulin, clone 1009, Cat. No. MABS2156, is a mouse monoclonal antibody that detects Thrombomodulin and has been tested for use in Flow Cytometry, Immunocytochemistry, Immunohistochemistry (Paraffin), Inhibition/Function Analysis, Radioimmunoassay, and Western Blotting.
More>>Anti-Thrombomodulin, clone 1009, Cat. No. MABS2156, is a mouse monoclonal antibody that detects Thrombomodulin and has been tested for use in Flow Cytometry, Immunocytochemistry, Immunohistochemistry (Paraffin), Inhibition/Function Analysis, Radioimmunoassay, and Western Blotting. Less<<
Empfohlene Produkte
Übersicht
Replacement Information
Description
Catalogue Number
MABS2156-100UL
Description
Anti-Thrombomodulin Antibody, clone 1009
Alternate Names
TM
Fetomodulin
CD141
Background Information
Thrombomodulin (UniProt: P07204; also known as TM, Fetomodulin, CD141) is encoded by the THBD (also known as THRM) gene (Gene ID: 7056) in human. Thrombomodulin is a single-pass type I membrane protein that is constitutively expressed on the surface of vascular endothelial cells. It serves as a high-affinity receptor for all thrombin forms and acts as a cofactor for the thrombin-dependent activation of protein C. It forms a 1:1 stoichiometric complex with thrombin. This complex is responsible for the conversion of protein C to the activated protein C, which then in combination with its cofactor protein S, acts as an anticoagulant by inactivating factors Va and VIIIa. Thrombomodulin is also reported to play a role in downregulating fibrinolysis and inflammation through activation of thrombin-activatable fibrinolysis inhibitor (TAFI). Thrombomodulin is synthesized with a signal peptide (aa 1-18), which is subsequently cleaved off to generate the mature form that contains a long extracellular domain (aa 19-515), a transmembrane domain (aa 516-539), and a cytoplasmic domain (540-575). Its extracellular region also contains a binding site (aa 481-515) for alpha-L/beta-2 and alpha-M/beta-2 integrin. (Ref.: Stearns-Kurosawa, DJ et al., (1996). Proc. Natl. Acad. Sci. USA Vol. 93(19); 10212-10216; Woodley-Cook, J et al. (2006). Mol. Cancer Ther. 5(12); 3303-3311).
References
Product Information
Format
Purified
Presentation
Purified mouse monoclonal antibody IgG1 in buffer containing 0.1 M Tris-Glycine (pH 7.4), 150 mM NaCl with 0.05% sodium azide.
Applications
Application
Anti-Thrombomodulin, clone 1009, Cat. No. MABS2156, is a mouse monoclonal antibody that detects Thrombomodulin and has been tested for use in Flow Cytometry, Immunocytochemistry, Immunohistochemistry (Paraffin), Inhibition/Function Analysis, Radioimmunoassay, and Western Blotting.
Key Applications
Flow Cytometry
Immunocytochemistry
Immunohistochemistry (Paraffin)
Inhibits Activity/Function
Radioimmunoassay
Western Blotting
Application Notes
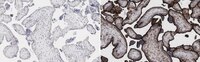
Immunohistochemistry (Paraffin) Analysis: A 1:50 dilution from a representative lot detected Thrombomodulin in human tonsil tissue sections.
ELISA Analysis: A representative lot detected Thrombomodulin in ELISA applicaitons (Woodley-Cook, J., et. al. (2006). Mol Cancer Ther. 5(12):3303-11).
Radioimmunoassay Analysis: A representative lot detected Thrombomodulin in Radioimmunoassay applications (Woodley-Cook, J., et. al. (2006). Mol Cancer Ther. 5(12):3303-11).
Inhibits Activity/Function Analysis: A representative lot of this blocked thrombin binding and reduced Protein C activation (Stearns-Kurosawa, D.J., et. al. (1996). Proc Natl Acad Sci USA. 93(19):10212-6).
Flow Cytometry Analysis: A representative lot detected Thrombomodulin in Flow Cytometry applications (Stearns-Kurosawa, D.J., et. al. (1996). Proc Natl Acad Sci USA. 93(19):10212-6; Woodley-Cook, J., et. al. (2006). Mol Cancer Ther. 5(12):3303-11).
Western Blotting Analysis: A representative lot detected Thrombomodulin in Western Blotting applications. (Xu, J., et. al. (1999). J Biol Chem. 274(10):6704-10; Woodley-Cook, J., et. al. (2006). Mol Cancer Ther. 5(12):3303-11).
Immunocytochemistry Analysis: A representative lot detected Thrombomodulin in Immunocytochemistry applications (Campbell, J.E., et. al. (2010). Blood. 116(26):6082-91).
Immunohistochemistry (Paraffin) Analysis: A representative lot detected Thrombomodulin in Immunohistochemistry applications (Trotman, W.E., et. al. (2011). Histochem Cell Biol. 135(2):141-52).
Biological Information
Immunogen
Recombinant fragment corresponding to 241 amino acids corresponding to six EGF repeats from human Thrombomodulin.
Clone
1009
Concentration
Please refer to lot specific datasheet.
Host
Mouse
Specificity
Clone 1009 is a mouse monoclonal antibody that detects Thrombomodulin.
Evaluated by Immunohistochemistry (Paraffin) in human placenta tissue sections.
Immunohistochemistry (Paraffin) Analysis: A 1:50 dilution of this antibody detected Thrombomodulin in human placenta tissue sections.
Usage Statement
Unless otherwise stated in our catalog or other company documentation accompanying the product(s), our products are intended for research use only and are not to be used for any other purpose, which includes but is not limited to, unauthorized commercial uses, in vitro diagnostic uses, ex vivo or in vivo therapeutic uses or any type of consumption or application to humans or animals.
The valve sinuses of the deep venous system are frequent sites of venous thrombus initiation. We previously reported that, in comparison with the non-valvular lumenal endothelium, the valve sinus endothelium had decreased expression of von Willebrand factor (vWF) and increased expression of endothelial protein C receptor (EPCR) and thrombomodulin (TM), suggesting alteration in the procoagulant/anticoagulant balance. We hypothesized that increased stasis in the deeper recesses of the venous valves would be associated with a gradient of increased thromboresistance. Expression of EPCR, TM, and vWF was analyzed via quantitative confocal immunofluorescence in residual saphenous veins collected following coronary artery bypass procedures. In agreement with our hypothesis, endothelial expression of vWF in the valve sinus decreased from the uppermost to the deepest region of the valve sinus. In contrast to our hypothesis, EPCR expression decreased from the uppermost to the deepest region of the valve sinus (p < 0.001) and TM expression remained unchanged throughout the valve sinus. Comparison of the non-valvular lumenal endothelium with the valve sinus endothelium demonstrated significantly decreased vWF expression (p < 0.001) in the valvular sinus consistent with our previous report; however, we did not observe statistically significant differences in EPCR or TM expression in this comparison. In addition, remarkable inter-individual variation in expression of these three proteins was also observed. These findings suggest that the genesis of these observations is more complex than predicted by our initial hypothesis, likely due, at least in part, to the complex rheology of the valvular sinus microenvironment.
We have adapted the corn-trypsin inhibitor whole-blood model to include EA.hy926 as an endothelium surrogate to evaluate the vascular modulation of blood coagulation initiated by relipidated recombinant tissue factor (rTf) and a cellular Tf surrogate, lipopolysaccharide (LPS)-stimulated THP1 cells (LPS-THP-1). Compared with bare tubes, EA.hy926 with rTf decreased the rate of thrombin formation, ITS accumulation, and the production of fibrinopeptide A. These phenomena occurred with increased rates of factor Va (fVa) inactivation by cleavages at R(506) and R(306). Thus, EA.hy926 provides thrombin-dependent protein C activation and APC fVa inactivation. Comparisons of rTf with LPS-THP-1 showed that the latter gave reduced rates for TAT formation but equivalent fibrinopeptide A, and fV activation/inactivation. In the presence of EA.hy926, the reverse was obtained; with the surrogate endothelium and LPS-THP-1 the rates of TAT generation, fibrinopeptide release, and fV activation were almost doubled, whereas cleavage at R(306) was equivalent. These observations suggest cooperativity between the 2 cell surrogates. These data suggest that the use of these 2 cell lines provides a reproducible quasi-endothelial quasi-inflammatory cytokine-stimulated monocyte system that provides a method to evaluate the variations in blood phenotype against the background of stable inflammatory cell activator and a stable vascular endothelial surrogate.
Effects of the chemotherapeutic agent doxorubicin on the protein C anticoagulant pathway. Woodley-Cook, J; Shin, LY; Swystun, L; Caruso, S; Beaudin, S; Liaw, PC Mol Cancer Ther
5
3303-11
2005
Although chemotherapy treatment is associated with an increased risk of thrombosis, the pathogenic mechanisms for the thrombogenic effect of chemotherapeutic drugs are poorly understood. We hypothesize that exposure of vascular endothelial cells to chemotherapeutic agents results in the loss of a thromboresistant phenotype. In this study, we examined the effects of the chemotherapeutic agent doxorubicin on the endothelium-based protein C anticoagulant pathway. The endothelial cell protein C receptor (EPCR) and thrombomodulin are two endothelial cell surface receptors required for the conversion of zymogen protein C to the anticoagulant enzyme activated protein C. Exposure of human umbilical vein endothelial cells (HUVEC) to doxorubicin resulted in a dose- and time-dependent decrease in cell surface EPCR levels. This decrease occurred as a result of receptor shedding as well as from a down-regulation in EPCR mRNA levels. In contrast, doxorubicin treatment of HUVECs resulted in a dose- and time-dependent increase in cell surface thrombomodulin attributed to an up-regulation of thrombomodulin mRNA levels. The net effect of the doxorubicin-induced changes in EPCR and thrombomodulin levels was a decrease in the capacity of HUVECs to convert protein C to activated protein C. Preliminary studies suggest that doxorubicin free radical metabolites mediate the doxorubicin-induced changes in EPCR expression but not those of thrombomodulin expression. In summary, these results suggest that doxorubicin alters the hemostatic balance of endothelial cells by down-regulating the endothelium-based protein C anticoagulant pathway.
Reconstitution of the human endothelial cell protein C receptor with thrombomodulin in phosphatidylcholine vesicles enhances protein C activation. Xu, J; Esmon, NL; Esmon, CT J Biol Chem
274
6704-10
1998
Blocking protein C binding to the endothelial cell protein C receptor (EPCR) on the endothelium is known to reduce protein C activation rates. Now we isolate human EPCR and thrombomodulin (TM) and reconstitute them into phosphatidylcholine vesicles. The EPCR increases protein C activation rates in a concentration-dependent fashion that does not saturate at 14 EPCR molecules/TM. Without EPCR, the protein C concentration dependence fits a single class of sites (Km = 2.17 +/- 0.13 microM). With EPCR, two classes of sites are apparent (Km = 20 +/- 15 nM and Km = 3.2 +/- 1.7 microM). Increasing the EPCR concentration at a constant TM concentration increases the percentage of high affinity sites. Holding the TM:EPCR ratio constant while decreasing the density of these proteins results in a decrease in the EPCR enhancement of protein C activation, suggesting that there is little affinity of the EPCR for TM. Negatively charged phospholipids also enhance protein C activation. EPCR acceleration of protein C activation is blocked by anti-EPCR antibodies, but not by annexin V, whereas the reverse is true with negatively charged phospholipids. Human umbilical cord endothelium expresses approximately 7 times more EPCR than TM. Anti-EPCR antibody reduces protein C activation rates 7-fold over these cells, whereas annexin V is ineffective, indicating that EPCR rather than negatively charged phospholipid provide the surface for protein C activation. EPCR expression varies dramatically among vascular beds. The present results indicate that the EPCR concentration will determine the effectiveness of the protein C activation complex.
The endothelial cell protein C receptor augments protein C activation by the thrombin-thrombomodulin complex. Stearns-Kurosawa, DJ; Kurosawa, S; Mollica, JS; Ferrell, GL; Esmon, CT Proc Natl Acad Sci U S A
93
10212-6
1996
Protein C activation on the surface of the endothelium is critical to the negative regulation of blood coagulation. We now demonstrate that monoclonal antibodies that block protein C binding to the endothelial cell protein C receptor (EPCR) reduce protein C activation rates by the thrombin-thrombomodulin complex on endothelium, but that antibodies that bind to EPCR without blocking protein C binding have no effect. The kinetic result of blocking the EPCR-protein C interaction is an increased apparent Km for the activation without altering the affinity of thrombin for thrombomodulin. Activation rates of the protein C derivative lacking the gamma-carboxyglutamic acid domain, which is required for binding to EPCR, are not altered by the anti-EPCR antibodies. These data indicate that the protein C activation complex involves protein C, thrombin, thrombomodulin, and EPCR. These observations open new questions about the control of coagulation reactions on vascular endothelium.