Wenn Sie das Fenster schließen, wird Ihre Konfiguration nicht gespeichert, es sei denn, Sie haben Ihren Artikel in die Bestellung aufgenommen oder zu Ihren Favoriten hinzugefügt.
Klicken Sie auf OK, um das MILLIPLEX® MAP-Tool zu schließen oder auf Abbrechen, um zu Ihrer Auswahl zurückzukehren.
Wählen Sie konfigurierbare Panels & Premixed-Kits - ODER - Kits für die zelluläre Signaltransduktion & MAPmates™
Konfigurieren Sie Ihre MILLIPLEX® MAP-Kits und lassen sich den Preis anzeigen.
Konfigurierbare Panels & Premixed-Kits
Unser breites Angebot enthält Multiplex-Panels, für die Sie die Analyten auswählen können, die am besten für Ihre Anwendung geeignet sind. Unter einem separaten Register können Sie das Premixed-Cytokin-Format oder ein Singleplex-Kit wählen.
Kits für die zelluläre Signaltransduktion & MAPmates™
Wählen Sie gebrauchsfertige Kits zur Erforschung gesamter Signalwege oder Prozesse. Oder konfigurieren Sie Ihre eigenen Kits mit Singleplex MAPmates™.
Die folgenden MAPmates™ sollten nicht zusammen analysiert werden: -MAPmates™, die einen unterschiedlichen Assaypuffer erfordern. -Phosphospezifische und MAPmate™ Gesamtkombinationen wie Gesamt-GSK3β und Gesamt-GSK3β (Ser 9). -PanTyr und locusspezifische MAPmates™, z.B. Phospho-EGF-Rezeptor und Phospho-STAT1 (Tyr701). -Mehr als 1 Phospho-MAPmate™ für ein einziges Target (Akt, STAT3). -GAPDH und β-Tubulin können nicht mit Kits oder MAPmates™, die panTyr enthalten, analysiert werden.
.
Bestellnummer
Bestellinformationen
St./Pkg.
Liste
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Wählen Sie bitte Spezies, Panelart, Kit oder Probenart
Um Ihr MILLIPLEX® MAP-Kit zu konfigurieren, wählen Sie zunächst eine Spezies, eine Panelart und/oder ein Kit.
Custom Premix Selecting "Custom Premix" option means that all of the beads you have chosen will be premixed in manufacturing before the kit is sent to you.
Catalogue Number
Ordering Description
Qty/Pack
List
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Spezies
Panelart
Gewähltes Kit
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
96-Well Plate
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
Weitere Reagenzien hinzufügen (MAPmates erfordern die Verwendung eines Puffer- und Detektionskits)
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
48-602MAG
Buffer Detection Kit for Magnetic Beads
1 Kit
Platzsparende Option Kunden, die mehrere Kits kaufen, können ihre Multiplex-Assaykomponenten in Kunststoffbeuteln anstelle von Packungen erhalten, um eine kompaktere Lagerung zu ermöglichen.
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Das Produkt wurde in Ihre Bestellung aufgenommen
Sie können nun ein weiteres Kit konfigurieren, ein Premixed-Kit wählen, zur Kasse gehen oder das Bestell-Tool schließen.
05-908
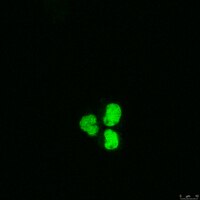
Sigma-AldrichAnti-RAD21 Antibody
Detect RAD21 with Anti-RAD21 Antibody (Mouse Monoclonal Antibody), that has been shown to work in IP, WB, ICC.
More>>Detect RAD21 with Anti-RAD21 Antibody (Mouse Monoclonal Antibody), that has been shown to work in IP, WB, ICC. Less<<
Anti-RAD21 Antibody: SDB (Sicherheitsdatenblätter), Analysenzertifikate und Qualitätszertifikate, Dossiers, Broschüren und andere verfügbare Dokumente.
Aka Scc1, part of the cohesin complex, Rad21 is a nuclear protein that associates with mitotic chromatin and is essential for the association of kinetochores with microtubules. Rad21/Scc1 s is cleaved by caspases, promoting apoptosis.
Background Information
The protein encoded by the Rad21 gene is highly similar to the gene product of Schizosaccharomyces pombe rad21, a gene involved in the repair of DNA double-strand breaks, as well as in chromatid cohesion during mitosis. This protein is a nuclear phospho-protein, which becomes hyperphosphorylated in cell cycle M phase. RAD21 also plays a role in apoptosis, via its cleavage by caspase 3 or caspase 7 during early steps of apoptosis: the C-terminal 64 kDa cleavage product may act as a nuclear signal to initiate cytoplasmic events involved in the apoptotic pathway. Recent work has shown that the RAD21 has a role in the invasion and metastasis of cancer cells
References
Product Information
Format
Purified
Control
HeLa nuclear extract
Presentation
Prtotein G purified antibody in 0.1M Tris-Glycine, 0.15M NaCl, 0.05% Sodium Azide, pH 7.4
The protein encoded by this gene is highly similar to the gene product of Schizosaccharomyces pombe rad21, a gene involved in the repair of DNA double-strand breaks, as well as in chromatid cohesion during mitosis. This protein is a nuclear phospho-protein, which becomes hyperphosphorylated in cell cycle M phase. The highly regulated association of this protein with mitotic chromatin specifically at the centromere region suggests its role in sister chromatid cohesion in mitotic cells.
FUNCTION: SwissProt: O60216 # Cleavable component of the cohesin complex, involved in chromosome cohesion during cell cycle, in DNA repair, and in apoptosis. The cohesin complex is required for the cohesion of sister chromatids after DNA replication. The cohesin complex apparently forms a large proteinaceous ring within which sister chromatids can be trapped. At metaphase-anaphase transition, this protein is cleaved by separase/ESPL1 and dissociates from chromatin, allowing sister chromatids to segregate. The cohesin complex may also play a role in spindle pole assembly during mitosis. Also plays a role in apoptosis, via its cleavage by caspase-3/CASP3 or caspase-7/CASP7 during early steps of apoptosis: the C-terminal 64 kDa cleavage product may act as a nuclear signal to initiate cytoplasmic events involved in the apoptotic pathway. SIZE: 631 amino acids; 71690 Da SUBUNIT: Cohesin complexes are composed of the SMC1 (SMC1A or SMC1B) and SMC3 heterodimer attached via their hinge domain, RAD21 which link them, and one STAG protein (STAG1, STAG2 or STAG3), which interacts with RAD21. Found in a complex with SMC1A, SMC3, CDCA5, PDS5A/APRIN and PDS5B/SCC-112. SUBCELLULAR LOCATION: Nucleus. Note=Associates with chromatin. Before prophase it is scattered along chromosome arms. During prophase, most of cohesin complexes dissociate from chromatin probably because of phosphorylation by PLK, except at centromeres, where cohesin complexes remain. At anaphase, it is cleaved by separase/ESPL1, leading to the dissociation of the complex from chromosomes, allowing chromosome separation. Once cleaved by caspase-3, the C-terminal 64 kDa cleavage product translocates to the cytoplasm, where it may trigger apoptosis. DOMAIN: SwissProt: O60216 The C-terminal part associates with the head of SMC1A, while the N-terminal part binds to the head of SMC3 (By similarity). PTM: Cleaved by separase/ESPL1 at the onset of anaphase. Cleaved by caspase-3 and caspase-7 at the beginning of apoptosis. The cleavage by ESPL1 and caspase-3 take place at different sites. & Phosphorylated; becomes hyperphosphorylated in M phase of cell cycle. The large dissociation of cohesin from chromosome arms during prophase may be partly due to its phosphorylation by PLK. SIMILARITY: Belongs to the rad21 family.
Molecular Weight
100-120kDa
Physicochemical Information
Dimensions
Materials Information
Toxicological Information
Safety Information according to GHS
Safety Information
Product Usage Statements
Quality Assurance
Routinely evaluated by immunoblot with HeLa nuclear extracts
Usage Statement
Unless otherwise stated in our catalog or other company documentation accompanying the product(s), our products are intended for research use only and are not to be used for any other purpose, which includes but is not limited to, unauthorized commercial uses, in vitro diagnostic uses, ex vivo or in vivo therapeutic uses or any type of consumption or application to humans or animals.
Architectural proteins CTCF and cohesin have distinct roles in modulating the higher order structure and expression of the CFTR locus. Gosalia, N; Neems, D; Kerschner, JL; Kosak, ST; Harris, A Nucleic acids research
42
9612-22
2014
Higher order chromatin structures across the genome are maintained in part by the architectural proteins CCCTC binding factor (CTCF) and the cohesin complex, which co-localize at many sites across the genome. Here, we examine the role of these proteins in mediating chromatin structure at the cystic fibrosis transmembrane conductance regulator (CFTR) gene. CFTR encompasses nearly 200 kb flanked by CTCF-binding enhancer-blocking insulator elements and is regulated by cell-type-specific intronic enhancers, which loop to the promoter in the active locus. SiRNA-mediated depletion of CTCF or the cohesin component, RAD21, showed that these two factors have distinct roles in regulating the higher order organization of CFTR. CTCF mediates the interactions between CTCF/cohesin binding sites, some of which have enhancer-blocking insulator activity. Cohesin shares this tethering role, but in addition stabilizes interactions between the promoter and cis-acting intronic elements including enhancers, which are also dependent on the forkhead box A1/A2 (FOXA1/A2) transcription factors (TFs). Disruption of the three-dimensional structure of the CFTR gene by depletion of CTCF or RAD21 increases gene expression, which is accompanied by alterations in histone modifications and TF occupancy across the locus, and causes internalization of the gene from the nuclear periphery.
Prophase pathway-dependent removal of cohesin from human chromosomes requires opening of the Smc3-Scc1 gate. Buheitel, J; Stemmann, O The EMBO journal
32
666-76
2013
Faithful transmission of chromosomes during eukaryotic cell division requires sister chromatids to be paired from their generation in S phase until their separation in M phase. Cohesion is mediated by the cohesin complex, whose Smc1, Smc3 and Scc1 subunits form a tripartite ring that entraps both DNA double strands. Whereas centromeric cohesin is removed in late metaphase by Scc1 cleavage, metazoan cohesin at chromosome arms is displaced already in prophase by proteolysis-independent signalling. Which of the three gates is triggered by the prophase pathway to open has remained enigmatic. Here, we show that displacement of human cohesin from early mitotic chromosomes requires dissociation of Smc3 from Scc1 but no opening of the other two gates. In contrast, loading of human cohesin onto chromatin in telophase occurs through the Smc1-Smc3 hinge. We propose that the use of differently regulated gates for loading and release facilitates unidirectionality of DNA's entry into and exit from the cohesin ring.
DNA double-strand breaks (DSBs) fuel cancer-driving chromosome translocations. Two related structural maintenance of chromosomes (Smc) complexes, cohesin and Smc5/6, promote DSB repair through sister chromatid homologous recombination (SCR). Here we show that the Smc5/6 subunit Mms21 sumoylates multiple lysines of the cohesin subunit Scc1. Mms21 promotes cohesin-dependent small ubiquitin-like modifier (SUMO) accumulation at laser-induced DNA damage sites in S/G2 human cells. Cells expressing the nonsumoylatable Scc1 mutant (15KR) maintain sister chromatid cohesion during mitosis but are defective in SCR and sensitive to ionizing radiation (IR). Scc1 15KR is recruited to DNA damage sites. Depletion of Wapl, a negative cohesin regulator, rescues SCR defects of Mms21-deficient or Scc1 15KR-expressing cells. Expression of the acetylation-mimicking Smc3 mutant does not bypass the requirement for Mms21 in SCR. We propose that Scc1 sumoylation by Mms21 promotes SCR by antagonizing Wapl at a step after cohesin loading at DSBs and in a way not solely dependent on Smc3 acetylation.
Cohesin regulates MHC class II genes through interactions with MHC class II insulators. Majumder, P; Boss, JM Journal of immunology (Baltimore, Md. : 1950)
187
4236-44
2010
Cohesin is a multiprotein, ringed complex that is most well-known for its role in stabilizing the association of sister chromatids between S phase and M. More recently, cohesin was found to be associated with transcriptional insulators, elements that are associated with the organization of chromatin into regulatory domains. The human MHC class II (MHC-II) locus contains 10 intergenic elements, termed MHC-II insulators, which bind the transcriptional insulator protein CCCTC-binding factor. MHC-II insulators interact with each other, forming a base architecture of discrete loops and potential regulatory domains. When MHC-II genes are expressed, their proximal promoter regulatory regions reorganize to the foci established by the interacting MHC-II insulators. MHC-II insulators also bind cohesin, but the functional role of cohesin in regulating this system is not known. In this article, we show that the binding of cohesin to MHC-II insulators occurred irrespective of MHC-II expression but was required for optimal expression of the HLA-DR and HLA-DQ genes. In a DNA-dependent manner, cohesin subunits interacted with CCCTC-binding factor and the MHC-II-specific transcription factors regulatory factor X and CIITA. Intriguingly, cohesin subunits were important for DNA looping interactions between the HLA-DRA promoter region and a 5' MHC-II insulator but were not required for interactions between the MHC-II insulators themselves. This latter observation introduces cohesin as a regulator of MHC-II expression by initiating or stabilizing MHC-II promoter regulatory element interactions with the MHC-II insulator elements, events that are required for maximal MHC-II transcription.
CTCF controls expression and chromatin architecture of the human major histocompatibility complex class II locus. Majumder, P; Boss, JM Molecular and cellular biology
30
4211-23
2009
The major histocompatibility complex class II (MHC-II) locus includes a dense cluster of genes that function to initiate immune responses. Expression of insulator CCCTC binding factor (CTCF) was found to be required for expression of all MHC class II genes associated with antigen presentation. Ten CTCF sites that divide the MHC-II locus into apparent evolutionary domains were identified. To define the role of CTCF in mediating regulation of the MHC II genes, chromatin conformation capture assays, which provide an architectural assessment of a locus, were conducted across the MHC-II region. Depending on whether MHC-II genes and the class II transactivator (CIITA) were being expressed, two CTCF-dependent chromatin architectural states, each with multiple configurations and interactions, were observed. These states included the ability to express MHC-II gene promoter regions to interact with nearby CTCF sites and CTCF sites to interact with each other. Thus, CTCF organizes the MHC-II locus into a novel basal architecture of interacting foci and loop structures that rearranges in the presence of CIITA. Disruption of the rearranged states eradicated expression, suggesting that the formation of these structures is key to coregulation of MHC-II genes and the locus.