Wenn Sie das Fenster schließen, wird Ihre Konfiguration nicht gespeichert, es sei denn, Sie haben Ihren Artikel in die Bestellung aufgenommen oder zu Ihren Favoriten hinzugefügt.
Klicken Sie auf OK, um das MILLIPLEX® MAP-Tool zu schließen oder auf Abbrechen, um zu Ihrer Auswahl zurückzukehren.
Wählen Sie konfigurierbare Panels & Premixed-Kits - ODER - Kits für die zelluläre Signaltransduktion & MAPmates™
Konfigurieren Sie Ihre MILLIPLEX® MAP-Kits und lassen sich den Preis anzeigen.
Konfigurierbare Panels & Premixed-Kits
Unser breites Angebot enthält Multiplex-Panels, für die Sie die Analyten auswählen können, die am besten für Ihre Anwendung geeignet sind. Unter einem separaten Register können Sie das Premixed-Cytokin-Format oder ein Singleplex-Kit wählen.
Kits für die zelluläre Signaltransduktion & MAPmates™
Wählen Sie gebrauchsfertige Kits zur Erforschung gesamter Signalwege oder Prozesse. Oder konfigurieren Sie Ihre eigenen Kits mit Singleplex MAPmates™.
Die folgenden MAPmates™ sollten nicht zusammen analysiert werden: -MAPmates™, die einen unterschiedlichen Assaypuffer erfordern. -Phosphospezifische und MAPmate™ Gesamtkombinationen wie Gesamt-GSK3β und Gesamt-GSK3β (Ser 9). -PanTyr und locusspezifische MAPmates™, z.B. Phospho-EGF-Rezeptor und Phospho-STAT1 (Tyr701). -Mehr als 1 Phospho-MAPmate™ für ein einziges Target (Akt, STAT3). -GAPDH und β-Tubulin können nicht mit Kits oder MAPmates™, die panTyr enthalten, analysiert werden.
.
Bestellnummer
Bestellinformationen
St./Pkg.
Liste
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Wählen Sie bitte Spezies, Panelart, Kit oder Probenart
Um Ihr MILLIPLEX® MAP-Kit zu konfigurieren, wählen Sie zunächst eine Spezies, eine Panelart und/oder ein Kit.
Custom Premix Selecting "Custom Premix" option means that all of the beads you have chosen will be premixed in manufacturing before the kit is sent to you.
Catalogue Number
Ordering Description
Qty/Pack
List
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Spezies
Panelart
Gewähltes Kit
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
96-Well Plate
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
Weitere Reagenzien hinzufügen (MAPmates erfordern die Verwendung eines Puffer- und Detektionskits)
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
48-602MAG
Buffer Detection Kit for Magnetic Beads
1 Kit
Platzsparende Option Kunden, die mehrere Kits kaufen, können ihre Multiplex-Assaykomponenten in Kunststoffbeuteln anstelle von Packungen erhalten, um eine kompaktere Lagerung zu ermöglichen.
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Das Produkt wurde in Ihre Bestellung aufgenommen
Sie können nun ein weiteres Kit konfigurieren, ein Premixed-Kit wählen, zur Kasse gehen oder das Bestell-Tool schließen.
Anti-phospho-PKR (Thr446) Antibody is an antibody against phospho-PKR (Thr446) for use in WB.
More>>Anti-phospho-PKR (Thr446) Antibody is an antibody against phospho-PKR (Thr446) for use in WB. Less<<
Anti-phospho-PKR (Thr446) Antibody: SDB (Sicherheitsdatenblätter), Analysenzertifikate und Qualitätszertifikate, Dossiers, Broschüren und andere verfügbare Dokumente.
FUNCTION: SwissProt: P19525 # Following activation by double-stranded RNA in the presence of ATP, the kinase becomes autophosphorylated and can catalyze the phosphorylation of the translation initiation factor EIF2S1, which leads to an inhibition of the initiation of protein synthesis. Double-stranded RNA is generated during the course of a viral infection. SIZE: 551 amino acids; 62094 Da SUBUNIT: Homodimer. Interacts with STRBP (By similarity). Interacts with DNAJC3. Inhibited by direct interaction with viral proteins such as HCV E2, HCV NS5A and influenza A NS1. Activated by the interaction with HIV-1 Tat. PTM: Autophosphorylated on several Ser and Thr residues. Autophosphorylation of Thr-451 is dependent on Thr-446 and is stimulated by dsRNA binding and dimerization. Autophosphorylation apparently leads to the activation of the kinase. SIMILARITY: SwissProt: P19525 ## Belongs to the protein kinase superfamily. Ser/Thr protein kinase family. GCN2 subfamily. & Contains 2 DRBM (double-stranded RNA-binding) domains. & Contains 1 protein kinase domain.
Molecular Weight
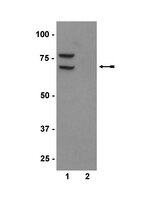
68 kDa
Physicochemical Information
Dimensions
Materials Information
Toxicological Information
Safety Information according to GHS
Safety Information
Product Usage Statements
Quality Assurance
routinely evaluated by immunoblot in RIPA lysates from HeLa cells that had been stimulated with IFN alpha overnight followed by treatment with calyculin A for 15 minutes
Usage Statement
Unless otherwise stated in our catalog or other company documentation accompanying the product(s), our products are intended for research use only and are not to be used for any other purpose, which includes but is not limited to, unauthorized commercial uses, in vitro diagnostic uses, ex vivo or in vivo therapeutic uses or any type of consumption or application to humans or animals.
Age-dependent decline in beta-cell proliferation restricts the capacity of beta-cell regeneration in mice. Tschen, SI; Dhawan, S; Gurlo, T; Bhushan, A Diabetes
58
1312-20
2009
The aim of this study was to elucidate whether age plays a role in the expansion or regeneration of beta-cell mass.We analyzed the capacity of beta-cell expansion in 1.5- and 8-month-old mice in response to a high-fat diet, after short-term treatment with the glucagon-like peptide 1 (GLP-1) analog exendin-4, or after streptozotocin (STZ) administration.Young mice responded to high-fat diet by increasing beta-cell mass and beta-cell proliferation and maintaining normoglycemia. Old mice, by contrast, did not display any increases in beta-cell mass or beta-cell proliferation in response to high-fat diet and became diabetic. To further assess the plasticity of beta-cell mass with respect to age, young and old mice were injected with a single dose of STZ, and beta-cell proliferation was analyzed to assess the regeneration of beta-cells. We observed a fourfold increase in beta-cell proliferation in young mice after STZ administration, whereas no changes in beta-cell proliferation were observed in older mice. The capacity to expand beta-cell mass in response to short-term treatment with the GLP-1 analog exendin-4 also declined with age. The ability of beta-cell mass to expand was correlated with higher levels of Bmi1, a polycomb group protein that is known to regulate the Ink4a locus, and decreased levels of p16(Ink4a)expression in the beta-cells. Young Bmi1(-/-) mice that prematurely upregulate p16(Ink4a)failed to expand beta-cell mass in response to exendin-4, indicating that p16(Ink4a)levels are a critical determinant of beta-cell mass expansion.beta-Cell proliferation and the capacity of beta-cells to regenerate declines with age and is regulated by the Bmi1/p16(Ink4a)pathway.
Translational inhibition of colonic epithelial heat shock proteins by IFN-gamma and TNF-alpha in intestinal inflammation. Shien Hu,Mae J Ciancio,Maor Lahav,Mikihiro Fujiya,Lev Lichtenstein,Shrikant Anant,Mark W Musch,Eugene B Chang Gastroenterology
133
2007
Inducible heat shock proteins (iHsp), Hsp25/27 and Hsp70, play essential roles in protecting cells against stress and, in intestinal mucosal inflammation, potentially lessening the extent and severity of injury. We examined the expression and regulation of iHsp in human and experimental inflammatory bowel diseases (IBD) and in vitro.
Upstream signaling pathways leading to the activation of double-stranded RNA-dependent serine/threonine protein kinase in beta-amyloid peptide neurotoxicity. Suen, Ka-Chun, et al. J. Biol. Chem., 278: 49819-27 (2003)
2003
One of the hallmarks of Alzheimer's disease is extracellular accumulation of senile plaques composed primarily of aggregated beta-amyloid (Abeta) peptide. Treatment of cultured neurons with Abeta peptide induces neuronal death in which apoptosis is suggested to be one of the mechanisms. We have demonstrated previously that Abeta peptide induces activation of double-stranded RNA-dependent serine/threonine protein kinase (PKR) and phosphorylation of eukaryotic initiation factor 2alpha (eIF2alpha) in neurons in vitro. Degenerating neurons in brain tissues from Alzheimer's disease patients also displayed high immunoreactivity for phosphorylated PKR and eIF2alpha. Our previous data have also indicated that PKR plays a significant role in mediating Abeta peptide-induced neuronal death, because neurons from PKR knockout mice and neuroblastoma SH-SY5Y cells stably transfected with dominant negative mutant of PKR are less susceptible to Abeta peptide toxicity. Therefore, it is important to understand how PKR is activated by Abeta peptide. We report here that inhibition of caspase-3 activity reduces phosphorylation of PKR and to a certain extent, cleavage of PKR and eIF2alpha in neurons exposed to Abeta peptide. Calcium release from the endoplasmic reticulum and activation of caspase-8 are the upstream signals modulating the caspase-3-mediated activation of PKR by Abeta peptide. Although in other systems HSP90 serves as a repressor for PKR, it is unlikely the candidate for caspase-3 to affect PKR activation in neurons after Abeta peptide exposure. Elucidation of the upstream pathways for PKR activation can help us to understand how this kinase participates in Abeta peptide neurotoxicity and to develop effective neuroprotective strategy.
Translation inhibition in apoptosis: caspase-dependent PKR activation and eIF2-alpha phosphorylation. Saelens, X, et al. J. Biol. Chem., 276: 41620-8 (2001)
2001
The protein kinase PKR is a major player in the cellular antiviral response, acting mainly by phosphorylation of the alpha-subunit of the eukaryotic translation initiation factor 2 (eIF2-alpha) to block de novo protein synthesis. PKR activation requires binding of double-stranded RNA or PACT/RAX proteins to its regulatory domain. Since several reports have demonstrated that translation is inhibited in apoptosis, we investigated whether PKR and eIF2-alpha phosphorylation contribute to this process. We show that PKR is proteolysed and that eIF2-alpha is phosphorylated at the early stages of apoptosis induced by various stimuli. Both events coincide with the onset of caspase activity and are prevented by caspase inhibitors. Using site-directed mutagenesis we show that PKR is specifically proteolysed at Asp(251) during cellular apoptosis. This site is cleaved in vitro by recombinant caspase-3, caspase-7, and caspase-8 and not by the proinflammatory caspase-1 and caspase-11. The released kinase domain efficiently phosphorylates eIF2-alpha at the cognate Ser(51) residue, and its overexpression in mammalian cells impairs the translation of its own mRNA and of reporter mRNAs. Our results demonstrate a new and caspase-dependent activation mode for PKR, leading to eIF2-alpha phosphorylation and translation inhibition in apoptosis.